Happy 2013, everyone!
Recently I announced a code package I was working on, called
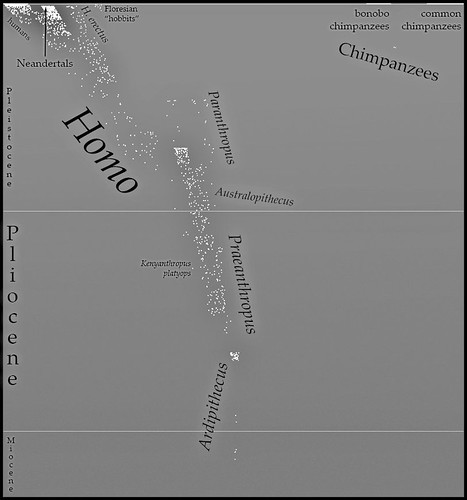
Haeckel, for generating vector-based charts related to evolutionary biology. Here's an image I've created using it:
 |
| Known Great Ape Individuals |
This chart represents all known hominid individuals (
Hominidae = great apes, including humans and stem-humans) from the Messinian to the present, erring on the conservative side when the material is too poor to determine the exact number.
If you've been following this blog for a few years you may remember
an earlier version of this. I've done a lot of refinement to the data since then. The earlier versions were dissatisfying to me because the horizontal axis was essentially arbitrary. For this version I used matrices from a phylogenetic analysis (
Strait and Grine 2004, Table 3 and Appendix C) of craniodental characters to generate a distance matrix, and then inferred positions for other taxa based on phylogenetic proximity and containing clade. This is similar to the metric I used in
this chart, except that it incorporates Appendix C, us
es inference, and averages distance from humans against distance from [Bornean] orangutans. Don't be mistaken — this is still arbitrary. But it's a bit closer to something real.
Stray notes:
- I'm pretty sure there are Pliocene stem-orangutans somewhere, right? Might have some work left to do on that data.
- The dot with no taxon above "Australopithecus" is an indeterminate stem-human from Laetoli. It should probably go further left.
- The Ardipithecus bubble includes the poorly-known "Australopithecus" praegens. (Although in some runs it moves outside — there's a random element to the plotting.)
- The Holocene is barely visible up at the top. What a worthless epoch.
- Homo floresiensis (hobbits) are far to the left of Homo sapiens because I placed them outside Clade(Homo erectus ∪ Homo sapiens).
- You may recall Lufengpithecus? wushanensis as "Wushan Man", as it was originally placed in Homo erectus. (Hey, it's just teeth.)
- A couple of fossil chimpanzees, lots of fossil orangutans, but no fossil gorillas. :(
- (Unless you count Chororapithecus, but that's pre-Messinian. Very pre-Messinian. Suspiciously pre-Messinian....)
- Look at all that overlap between Homo, Paranthropus, and Australopithecus!
- I have a feeling, though, that if I added another dimension, Paranthropus and Homo would jut out in opposite directions.
- Reclassifying Australopithecus sediba as Homo sediba would also decrease the overlap. (Although its position is inferred — actually scoring it might do the same thing.)
- It's frustrating that the type species of Australopithecus and Paranthropus are also just about the most similar species across the two genera.
- Kenyanthropus and Praeanthropus have been provisionally sunk into Australopithecus.
- Should we just sink Orrorin and Sahelanthropus into Ardipithecus? Why not?
- My guess is that if I added postcranial characters, the stem-humans would all shift right (humanward). Oh, for a good matrix of postcranial characters....
Update
Oh yeah, and if you want a peek at the data,
go here.